Bičíky a další povrchové struktury
| Website: | Moodle Veterinární univerzita Brno |
| Kurs: | Mikrobiologie potravin a mikrobiologické laboratorní metody 1 |
| Buch: | Bičíky a další povrchové struktury |
| Gedruckt von: | Gast |
| Datum: | Montag, 2. Juni 2025, 14:30 |
Beschreibung
Kapitola uvádí informace a stavbě bakterálních bičíků a dalších povrchových struktur.
Orgány pohybu - bičíky
Pohyb bakterií je umožněn díky bičíkům. Bičíky jsou tenká vlákna mnohonásobně delší než bakteriální buňka (tloušťka 20 – 30 nm, délka asi 20 µm).
Podle počtu a umístění bičíků rozlišujeme několik typů bakterií. Jedná se o atricha (zcela bez bičíků), monotricha (jediný bičík umístěný na pólu buňky), lofotricha (svazek bičíků umístěný na jednom pólu buňky), amfitricha (svazek bičíků umístěný na obou pólech buňky) a peritricha (bičíky pokrývají celou buňku).
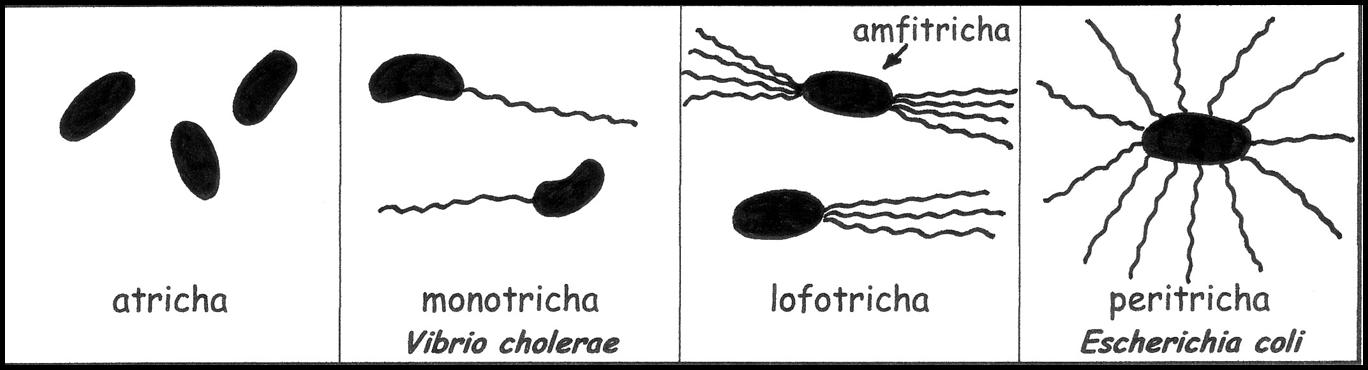
I přes svoji délku, je tloušťka bičíků tak nepatrná, že jejich pozorování světelným mikroskopem je velmi obtížné. Pro jejich zviditelnění se používají náročné barvicí postupy, např. stříbření. Běžně můžeme bičíky pozorovat v elektronovém mikroskopu. Jednoduchým způsobem průkazu pohyblivosti bakterií je jejich kultivace v polotuhých živných půdách.
Stavba bičíku
Každý bičík se skládá ze 3 částí – vlákna bičíku, háčku a bazální části (osa a kruhové destičky).
Vlákno bičíku je složeno z bílkoviny flagelinu. Aminokyselinové složení flagelinu (a tím i jeho antigennost) se u jednotlivých druhů či serotypů bakterií liší, hovoříme o tzv. bičíkových H-antigenech. Molekuly flagelinu jsou uspořádány ve šroubovici, uvnitř je vlákno duté. Po své syntéze v cytoplasmě, prochází molekuly flagelinu dutinou uprostřed vlákna bičíku až na jeho rostoucí konec, kde se díky svému tvaru automaticky zapojují mezi prostorově komplementární sousední molekuly.
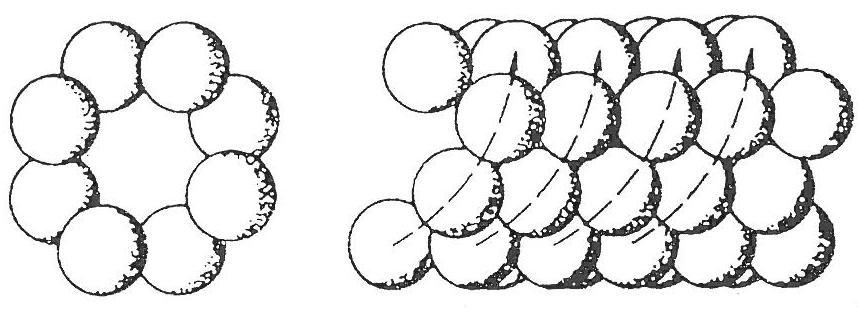
Uspořádání molekul flagelinu
(zdroj: KAPRÁLEK, F. Základy bakteriologie. 1. vyd. Praha, ČR: Univerzita Karlova v Praze, Nakladatelství Karolinum. 2000. 241 p.)
Háček je silnější část zpevňující spodní konec vlákna. Vytváří flexibilní spojení mezi fixovanou bazální částí a rigidním (neohebným) vláknem. Háček je také tvořen z bílkovinných podjednotek, které jsou ale odlišné od flagelinu.
Bazální část kotví bičík v buněčné stěně a cytoplasmatické membráně. U gramnegativních bakterií se skládá ze dvou párů kruhových destiček (P-, L-, S- a M-kruh), jejichž středem prochází osa připojující je k háčku. Grampozitivní bakterie mají pouze spodní dvojici destiček (S- a M-kruh). Spodní prstenec, tedy M-kruh, je v obou případech zakotven v cytoplasmatické membráně, ostatní prstence potom v dalších částech buněčné stěny.
Bičík gramnegativních bakterií
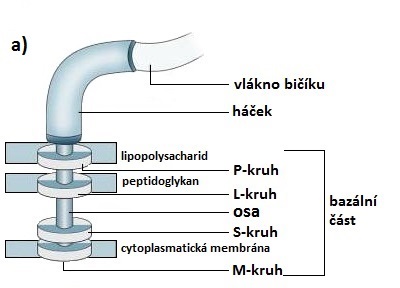
(upraveno podle: KUMAR, S. Textbook of Microbiology. 1st ed. New Delhi, India: Jaypee Brothers Medical Publisher (P) Ltd. 2012. 784 p.)
Bičík grampozitivních bakterií
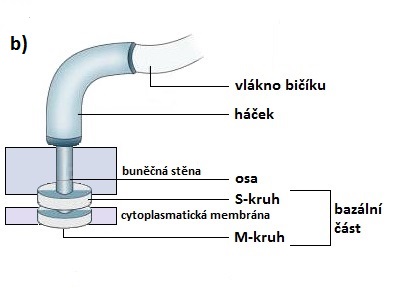
(upraveno podle: KUMAR, S. Textbook of Microbiology. 1st ed. New Delhi, India: Jaypee Brothers Medical Publisher (P) Ltd. 2012. 784 p.)
Pohyb bičíku
Bičík rotuje na způsob lodního šroubu, kdy volně otáčivý M-kruh funguje z mechanického hlediska jako rotor a nepohyblivá sousední destička (S-kruh) funguje jako stator. Zdrojem energie pro rotaci je elektrochemický potenciál (gradient protonů) na cytoplasmatické membráně. Bakterie má možnost měnit rychlost i směr otáčení bičíku.
Rychlost pohybu bakterie je poměrně velká, až 50 µm za sekundu. Při pohybu bakterie se střídají fáze přímočarého pohybu v náhodném směru s okamžiky, kdy se bakterie zastaví a točí se na místě („kotrmelcování“), potom následuje přímočarý pohyb v novém směru. Pohyb bakterií může být zcela náhodný nebo je ovlivněn přítomností určitých látek v prostředí a bakterie se pohybují do oblasti optimální koncentrace těchto látek – pozitivní či negativní chemotaxe. Uplatňují se zde chemotaktické receptory v cytoplasmatické membráně. Podobná reakce aerobních či anaerobních bakterií na kyslík se nazývá aerotaxe, reakce fototrofních bakterií na světlo potom fototaxe. Má-li buňka více bičíků, musí být jejich pohyb koordinován. Bylo prokázáno, že peritrichní bičíky při svém pohybu vytvářejí jediný svazek, který pohání buňku dopředu.
Orgánem pohybu spirochet jsou tzv. axiální vlákna. Jejich stavba je podobná jako u bičíků, vycházejí z pólů buňky, ale jsou uložena pod jejím povrchem. Svým zkracováním či prodlužováním uvádějí buňku do pohybu, který může být rotační, vývrtkovitý nebo hadovitý.
Další povrchové struktury
Pili (fimbrie)
Na povrchu gramnegativních bakterií můžeme pozorovat četná velmi křehká, krátká, rovná, dutá vlákna trčící všemi směry. Jedná se o fimbrie neboli pili (j.č. pilus). Na jedné buňce jich může být až několik set. Fimbrie jsou tvořeny proteinovými podjednotkami (tzv. piliny), pozorovat je můžeme v elektronovém mikroskopu. Mají antigenní vlastnosti. Jejich tvorba je ovlivněna podmínkami vnějšího prostředí (pH, teplota, obsah kyslíku).
Dělíme je na několik typů. Většina z nich má adhezivní funkci – usnadňují specifickou kolonizaci hostitele. Zvláštním typem jsou fimbrie kódované konjugativním plasmidem (tzv. F pili nebo též méně správně sex pili), které jsou poměrně velké a ohebné. Z bakterie označované jako buňka donorová vystupuje obvykle jeden sex pilus. Ten při konjugaci vytváří dutý můstek mezi donorovou a recipientní buňkou, kterým prochází plasmidová DNA z jedné buňky do druhé.
Curli (z angl. curl, kadeř, kudrna) jsou shluky štíhlých zprohýbaných vlákének na povrchu některých escherichií a salmonel, které na sebe váží sérové bílkoviny (fibronektin, bílkoviny nezbytné pro srážení krve) a mohly by odpovídat za některé příznaky sepse.
Další povrchové struktury
Extracelulární polymery
Některé grampozitivní i gramnegativní bakterie vytváří nad buněčnou stěnou další polysacharidovou vrstvu různé tloušťky a stavby. Tvorba extracelulárních polymerů není pozorována u všech bakterií, je mimo jiné ovlivněna i složením vnějšího prostředí.
Je-li tato vrstva kondenzovaná a ostře ohraničená, označuje se jako pouzdro – kapsule. Pouzdro má antigenní vlastnosti (kapsulární K antigen u Escherichia coli či Klebsiella pneumoniae) a přispívá k virulenci a invazivitě patogenních bakterií (ochrana před napadením bakteriofágy, proti účinku protilátek či zvýšení odolnosti vůči fagocytóze). Pouzdro Bacillus anthracis je jako jediné tvořeno polypeptidy, konkrétně poly-D-glutamovou kyselinou.
Polysacharidová vrstva může být i ve formě volného slizu (např. u Leuconostoc mesenteroides – rosolovatění slazených minerálních vod v lahvích, Bacillus spp. – nitkovitost pečiva nebo řídké síťoviny polysacharidových vláken – tzv. glykokalyx, která hraje klíčovou roli v adherenci bakterií na různé povrchy (zubní plak, tvorba biofilmů).
Extracelulární polymery nepřijímají běžná barviva, dají se nejlépe znázornit negativním barvením např. pomocí tuše a karbolfuchsinu. Částečky tuše polysacharidovou vrstvou neproniknou, takže ve výsledku pozorujeme na šedočerném pozadí červeně obarvené buňky obklopené prázdným dvorcem.
Další povrchové struktury
S-vrstva
Další objevenou povrchovou strukturou je tenká vrstva strukturovaného proteinu či glykoproteinu, která se označuje jako S-vrstva. Jedná se o pravidelnou, plochou, dvojrozměrnou vrstvu monomolekulárních bílkovinných podjednotek tvořících na povrchu buňky určitou síťovinu či „krystalickou mřížku“. Druhově specifické bílkovinné podjednotky mohou mít např. tvar čtverce nebo šestiúhelníku, často vypadají jako dlaždice.